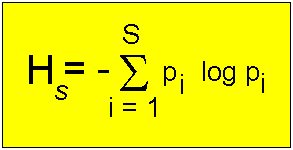Als ein Maß für die Biodiversität, bzw. Mannigfaltigkeit der Arten wird häufig der Shannon-Wiener-Index
Hs verwendet, welcher oft fälschlicherweise als Shannon-Weaver-Index bezeichnet wird,
da Shannon seinen Beitrag zusammen mit einem weiteren Artikel von Weaver in Buchform publizierte.
Wiener und Shannon entwickelten unabhängig voneinander die Grundlage dieser Funktion, und daher ist sie nach
beiden benannt (siehe: Washington, H.G. 1984: Diversity, biotic and similarity indices - a review with special relevance to aquatic
ecosystems. Water Resources, 18; p. 665, 653-694).
Allgemeines
Die Vielfalt kann man zunächst vereinfacht als die Anzahl der Arten definieren.
Das mag für einige Untersuchungen ein brauchbarer Wert sein, für die meisten Situationen ist er jedoch nicht ideal.
Oft reicht es nicht aus, die Anzahl der Arten zu wissen.
Von Bedeutung ist auch die relative Häufigkeit oder Dominanz jeder Art.
Betrachten wir zunächst die Überlebenschancen einer Art: Wenn ein Räuber von 10 Beutearten abhängig ist,
die alle über einen lange Zeitraum hinweg ungefähr die gleiche Abundanz haben,
so haben alle 10 Arten für die Ernährung des Räubers eine vergleichbare Bedeutung.
Falls nur eine Art sehr häufig und neun sehr selten sind, dann spielen die neun Arten mit nur wenigen Individuen eine unbedeutende
Rolle bei der Ernährung und Regulierung der Räuberpopulation.
Sie verhält sich im wesentlichen so, als ob nur eine einzige Art als Beute zur Verfügung stünde.
In diesem Fall ist das Risiko zu verhungern für den Räuber sehr groß, wenn diese eine Hauptbeuteart nicht mehr verfügbar ist.
Das Überleben des Beutegreifers ist somit nicht gesichert, sondern gefährdet und instabil.
Demnach muß ein Maß für die Mannigfaltigkeit bzw. Diversität auch die relativen Häufigkeiten (=Dominanz) der Arten berücksichtigen.
Mehrere Maße sind vorgeschlagen worden, u. a. der Simpson-Index.
Der am häufigsten benutzte Index wird teilweise als das Maß für die Entropie oder als der
Shannon-Wiener-Informationsgehalt oder einfach als der Informationsgehalt des Systems bezeichnet:
Die Shannon-Wiener-Formel
Hs = das Symbol für das Ausmaß der Mannigfaltigkeit (Heterogenität)
in einer Gruppe von s Arten.
Die Klassifizierungskategorie ist in diesem Fall die Art (spezies, daher der Index s),
aber es können auch andere Kategorien (z.B. genetische Variabilität oder funktionale Gruppen) benutzt werden. s = Anzahl der vorhandenen Arten; pi = die relative Abundanz der i-ten Art, gemessen von 0,0 bis 1,0
(wenn z.B. die betrachtete Art die zweithäufigste ist, so ist i=2;
und wenn die Dominanz dieser Art 10% beträgt, bzw. 10% aller Organismen dieser Art angehören,
so ist pi =0,10; log pi = der Logarithmus von pi;
er kann ausgedrückt werden in bezug auf die Basis 2, e oder 10.
Der Einfachheit halber wird hier die Basis e des natürlichen Logarithmus verwendet.
Da pi
stets zwischen 0 und 1,0 liegt, sind alle Logarithmen negativ.
Deshalb wird ein Minuszeichen hinzugefügt, damit Hs
positiv wird. Dieser Maßstab für die Mannigfaltigkeit der Arten in einem Areal wird vor allem deswegen verwendet,
weil er dem Entropiemaß in der Thermodynamik und dem Informationsgehalt in der Informationstheorie entspricht.
In allen drei Wissenschaftszweigen - Physik, Informationstheorie und Ökologie - gibt Hs
den Grad der 'Ungewißheit' an. Ein größeres Hs
bedeutet: Beim zufälligen Herausgreifen eines Atoms oder einer Nachricht oder eines einzelnen Organismus können wir nicht
sicher sein, welche Art Atom, Nachricht oder Organismus wir erhalten. Ist in einer Gruppe z.B. nur eine einzige Art
vertreten, so ist die Ungewißheit über den blindlings herausgegriffenen Organismus gleich Null. Entsprechend wird auch der
Wert Hs gleich Null sein,
da für nur eine Art pi =1
und der Ausdruck log pi = log1 = 0 ist.
Beispiel
Berechnung der Mannigfaltigkeit Hs
bezüglich der Verteilung von Mäusen auf zwei Probeflächen (Forstkulturen).
Erste Probefläche:
Art
Anzahl
i
pi
log pi
pi*log pi
Erdmaus
10
1
0,25
-1,386
-0,34657
Feldmaus
10
2
0,25
-1,386
-0,34657
Rötelmaus
10
3
0,25
-1,386
-0,3465
Waldmaus
10
4
0,25
-1,386
-0,34657
Summe:
40
Summe: -1,3863
Hs = 1,3863
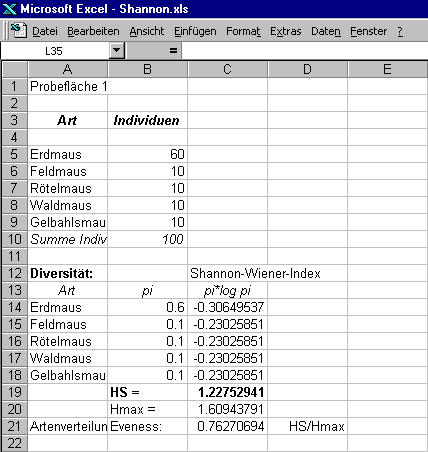
Zweite Probefläche:
Art
Anzahl
i
pi
log pi
pi*log pi
Erdmaus
60
1
0,6
-0,511
-0,30649
Feldmaus
10
2
0,1
-2,303
-0,23026
Rötelmaus
10
3
0,1
-2,303
-0,23026
Waldmaus
10
4
0,1
-2,303
-0,23026
Gelbhalsmaus
10
5
0,1
-2,303
-0,23026
Summe:
100
Summe: -1,2275
Hs = 1,2275
Obwohl auf der zweiten Probefläche mehr Mäusearten vorkommen als auf der ersten,
ist der errechnete Shannon-Wiener-Index Hs
für die zweite Fläche etwas geringer. Dies ist auf die unausgeglichenen Dominanzverhältnisse, bzw. die sehr
unterschiedlichen Artenanteile auf der zweiten Fläche zurückzuführen.
Das Entropiemaß der Mannigfaltigkeit (Diversität) hat verschiedene Vorteile. Es hat beispielsweise keine obere Grenze.
Werden zu einer Artengruppe noch weitere Arten hinzugefügt, so kann Hs
unbegrenzt zunehmen; eine unendliche Anzahl von Arten würde theoretisch zu einem unendlich
großen Hs führen.
Ebenso können die für jede Spezies errechneten Entropiemaße einfach addiert werden.
Sie ergeben dann den Gesamtwert der Mannigfaltigkeit für alle Arten.
Der Index Hs ist außerdem unabhängig von der Abundanz,
bzw. der Populationsgröße. Wenn z.B. im oben gezeigten Beispiel auf der zweiten Probefläche anstelle der
Individuenzahlen 60,10,10,10,10 folgende Werte eingetragen werden:
6,1,1,1,1 so ergibt sich der gleiche Diversitätsindex 1,2275.
Für jede gegebene Anzahl von Arten wird Hs
am größten sein (Hs=Hmax),
wenn die Arten alle gleich häufig sind (p1= p2=
p3= p4 .....). Das ist auch intuitiv richtig,
denn die Ungewißheit über das Vorkommen einer bestimmten Spezies ist in der Tat am größten,
wenn alle Arten, zu denen diese Spezies möglicherweise gehören kann,
mit der gleichen Wahrscheinlichkeit vertreten sind. Hmax
kann allerdings einfacher mit folgender Formel ermittelt werden:
Hmax = -log(1/i); i = Anzahl der Arten;
Im oben gezeigten Beispiel (Probefläche 2) beträgt Hmax
= -log(1/5)=1.60943791.
Den gleichen Wert erhält man, wenn man in die Tabelle anstelle der
Individuenzahlen 60,10,10,10,10 die Werte 20,20,20,20,20 oder
1,1,1,1,1 einträgt und den Diversitätsindex Hs
mit der Shannon-Wiener-Formel berechnet.
Das bedeutet: In die Formel kann man sowohl die absoluten Individuenzahlen jeder Art einsetzen,
als auch zuverlässige Index- oder Stichprobenwerte, bzw. relative Abundanzwerte (z.B. Prozentwerte),
wenn man die absoluten Werte nicht ermitteln kann.
Das Verhältnis von Hs/Hmax
wird häufig als Eveness E bezeichnet.
Dieser Quotient liegt zwischen den Werten 0 bis 1. Je stärker E von 1 verschieden ist,
desto mehr weichen die Dominanzverhältnisse verschiedener Arten von der Gleichverteilung ab.
Vollkommene Eveness (E=1) wird erreicht,
wenn alle Arten mit dem gleichen Anteil vertreten sind (Hs = Hmax).
Die verschiedenen Individuen kann man nicht nur nach Arten, sondern auch nach anderen Kriterien ordnen,
beispielsweise nach Nahrungsgilden oder bevorzugten Habitaten.
Für jede neue Klassifikation kann man einen eigenen Diversitätsindex Hs
und entsprechende Eveness E berechnen.
Man kann auch erkennen, welchen Beitrag eine einzelne Art, bzw. eine bestimmte Gilde oder Kategorie zur Gesamtdiversität leistet,
da jede Kategorie mit einem Anteil von pi*log pi
an der gesamten Mannigfaltigkeit der Kategorien (Arten) beteiligt ist.
Auf der Programm-CD ist in der Datei Shannon.bas
ein einfacher Basic-Quellcode enthalten, mit dem man den Shannon-Wiener-Diversitätsindex
Hs
und Hmax, sowie die Eveness E
beispielsweise in einer Excel-Tabelle selbst berechnen können.
Kopieren Sie diesen Quellcode und fügen Sie ihn als Makro in eine Excel-Arbeitsblatt ein.
Verwenden Sie für diesen Zweck das Excel-Menü:
Extras - Makro - Erstellen, bzw.
Extras - Makro - Bearbeiten.
Wenn Sie das Makro ausführen mit dem Excel -Menü Extras - Makro - Ausführen,
werden diese Parameter der Mannigfaltigkeit berechnet und in die Tabelle eingetragen (s. Abbildung oben).
Im Prinzip werden die gleichen Algorithmen auch im Programm MAPPER verwendet.
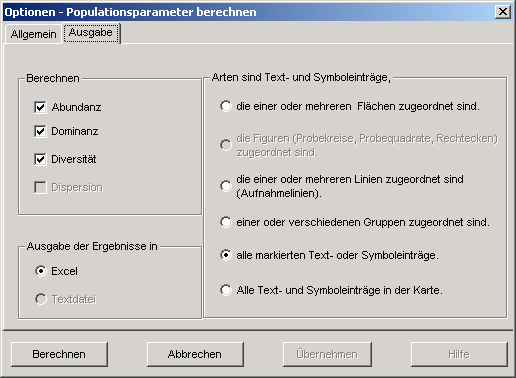
Wenn Sie in diesem Programm für faunistische Aufnahmen das nebenstehende Menü Berechnen
- Populationsparameter wählen und dann die Option Shannon-Wiener aktivieren,
schreibt MAPPER die Karteneinträge (Objekte) in eine Excel-Tabelle und trägt hier auch die errechneten Diversitätswerte ein.
Sie können dabei bestimmen, welche Karteneinträge Arten repräsentieren.
Beispielsweise können sämtliche Text- und Symboleinträge in einer Karte als Arten deklariert werden. Sie können auch
festlegen, dass nur die markierten oder die zu einem oder mehreren Polygonen (Flächen, Linien, Figuren) zugeordneten Text-
und Symboleinträge Arten bezeichnen.
Bedeutung der Biodiversität
Biodiversität wird häufig im Zusammenhang mit der Stabilität von Ökosystemen diskutiert.
In gestörten Systemen nimmt im allgemeinen die Mannigfaltigkeit (Biodiversität) ab. Anstelle der Artenvielfalt tritt dann die Einfalt
bzw. Artenarmut. Dabei ist meistens nicht nur die Zahl der Arten reduziert, sondern auch die Verteilung der restlichen Arten
pi zunehmend ungleich. In extrem einförmigen Systemen, wie
z.B. in Monokulturen dominiert häufig nur noch eine Art beispielsweise mit einer Dominanz von mehr als 80% aller Individuen
(pi = 0,8). Eine derart dominierende Art ist dann häufig ein phytophager Schädling,
wie z.B. der Kartoffelkäfer, der sich in einem Kartoffelfeld ohne Konkurrenten und Gegenspieler optimal vermehren kann.
Je größer und je einförmiger das betreffende Feld ist und je häufiger Störungen z.B. infolge von Bodenbearbeitung, Düngung
oder Pestizidanwendung eintreten, desto geringer wird die Biodiversität. Das gesamte System wird störungsanfälliger
und in zunehmendem Maße von Schadorganismen bedroht.
Naturnahe Systeme wie z.B. Urwälder haben dagegen eine sehr große Biodiversität.
Auch dort gibt es viele 'Schadorganismen'. Aber sie fallen in solchen sehr komplexen Systemen nicht ins Gewicht,
da sie aufgrund der hohen Biodiversität von vielen Konkurrenten und Gegenspielern in Schach gehalten werden.
Man kann deshalb sagen: In naturnahen Systemen mit einer großen Mannigfaltigkeit haben Schadorganismen keinen großen Einfluß.
Solche Systeme sind deshalb stabiler als die einförmigen, künstlichen oder gestützten Systeme, die der Mensch geschaffen hat.
Die tropischen Regenwälder haben beispielsweise viele Millionen Jahre ohne Zutun des Menschen überlebt.
Die intensiv bewirtschaftete Kulturlandschaft würde dagegen ohne den Einfluß des Menschen sehr rasch verschwinden.
Sie ist aufgrund der geringeren Artenvielfalt nicht alleine überlebensfähig, sondern abhängig von der Pflege und den steuernden Eingriffen
des Menschen.
Allerdings darf man die Bedeutung der Indexwerte auch nicht überbewerten. Viel entscheidender für die Stabilität vieler
Systeme ist nicht allein die Artenzahl bzw. ein Diversitätsindex, sondern das Beziehungsgeflecht der Arten im System. Dazu ein Beispiel:
Bei einer Massenvermehrung des Eichenwicklers treten meistes viele andere Insektenarten als Begleitarten auf.
In diesem Fall nimmt also die Artenvielfalt zu.
Entscheidend ist dabei, ob die im Verbund mit dem Eichenwickler auftretenden Arten ebenfalls grünes Eichenlaub vertilgen
und den Bestand des befallenen Eichenwalds gefährden oder
als Antagonisten die Zahl der Eichenwickler reduzieren und so mit negativer Rückkopplung und Redundanz dazu beitragen,
das System zu stabilisieren. Wenn man die Stabilität eines Systems verstehen und bewerten will,
sind eingehende Untersuchungen der komplexen Beziehungen im System unerlässlich,
wobei auch abiotische Einflüsse, z.B. Dürre, Überschwemmungen, Boden, Geologie und Eingriffe des Menschen zu berücksichtigen sind.
Ein Indexwert der Biodiversität ist dafür alleine nicht ausreichend.